
 Grunnen til at de fleste nukleotider (livets 'bokstaver': A, T, C og G) ikke er selekterbare, er p.g.a. lave kvotienter mellom signal og støy. Det samme gjelder hvorfor vi ikke kan selektere for mange nukleotider (baser) samtidig, er på grunn av raskt synkende kvotienter mellom signal og støy. En av hovedårsakene til at vi ikke kan redde genomet fra mutasjoner, er på grunn av allestedsnærværende støy. Når vi anvender seleksjon i forhold til hele genomet, synker kvotienten signal/støy raskt mot 0. Dette er en viktig grunn til at seleksjon kan virke på gennivå, men feiler på genomnivå.
Grunnen til at de fleste nukleotider (livets 'bokstaver': A, T, C og G) ikke er selekterbare, er p.g.a. lave kvotienter mellom signal og støy. Det samme gjelder hvorfor vi ikke kan selektere for mange nukleotider (baser) samtidig, er på grunn av raskt synkende kvotienter mellom signal og støy. En av hovedårsakene til at vi ikke kan redde genomet fra mutasjoner, er på grunn av allestedsnærværende støy. Når vi anvender seleksjon i forhold til hele genomet, synker kvotienten signal/støy raskt mot 0. Dette er en viktig grunn til at seleksjon kan virke på gennivå, men feiler på genomnivå. innebærer det 100% arvelighet (eks. blodtype). Om arveligheten er 0 for et trekk, bestemmes det kun ut fra omgivelsene (eks. tatovering). En egenskap som høyde er vanligvis høyst arvelig (h=0,3). Det innebærer at 30% av fenotype variasjonen er arvelig (selekterbar) variasjon. For meget komplekse trekk som fitness ('survival of fittest') er arveligheten meget lav. Kimura anslår den så lavt som 0,004 (Kimura, 1983, s30-31). Det skyldes at fitnes integrerer alle støytyper fra alle de ulike aspektene til individet.
innebærer det 100% arvelighet (eks. blodtype). Om arveligheten er 0 for et trekk, bestemmes det kun ut fra omgivelsene (eks. tatovering). En egenskap som høyde er vanligvis høyst arvelig (h=0,3). Det innebærer at 30% av fenotype variasjonen er arvelig (selekterbar) variasjon. For meget komplekse trekk som fitness ('survival of fittest') er arveligheten meget lav. Kimura anslår den så lavt som 0,004 (Kimura, 1983, s30-31). Det skyldes at fitnes integrerer alle støytyper fra alle de ulike aspektene til individet.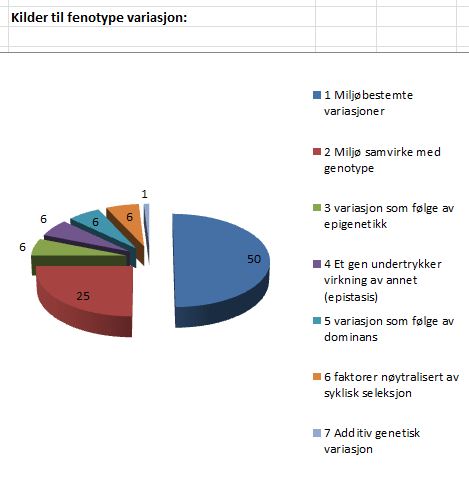 omgivelsene, kommer denne type av variasjon i tillegg til støy og interagerer med seleksjon. Denne andelen er vist i seksjon 2 kake-diagrammet.
omgivelsene, kommer denne type av variasjon i tillegg til støy og interagerer med seleksjon. Denne andelen er vist i seksjon 2 kake-diagrammet. Alternativt kan en ha en mengde froske-egg i en elv. Et stort antall av dem spises av fisk, før de klekkes. En stor mengde rumpetroll, plukkes av fugler, en båt kjører gjennom stimen og dreper noen hundre til etc: nesten all eliminasjon er tilfeldig. Vi ser igjen overlevelse av de heldigste. Den systematiske sortering etter fenotype, er hovedsakelig fraværende. Støy påvirker til de grader reproduksjons-suksess, at det ikke er aktuelt med terskel-verdier for slike seleksjons-scenarier. Det indikerer en lav korrelasjons-koeffisient, for at overlegen fenotype-kvalitet vil reprodusere. Kanskje 50% av reproduksjon-fiasko, er uavhengig av fenotype-kvalitet.
Alternativt kan en ha en mengde froske-egg i en elv. Et stort antall av dem spises av fisk, før de klekkes. En stor mengde rumpetroll, plukkes av fugler, en båt kjører gjennom stimen og dreper noen hundre til etc: nesten all eliminasjon er tilfeldig. Vi ser igjen overlevelse av de heldigste. Den systematiske sortering etter fenotype, er hovedsakelig fraværende. Støy påvirker til de grader reproduksjons-suksess, at det ikke er aktuelt med terskel-verdier for slike seleksjons-scenarier. Det indikerer en lav korrelasjons-koeffisient, for at overlegen fenotype-kvalitet vil reprodusere. Kanskje 50% av reproduksjon-fiasko, er uavhengig av fenotype-kvalitet. Tilfeldig genetisk drift er meget sterk, og kan overstyre effekten av endog viktige mutasjoner i små populasjoner. I små populasjoner er naturlig utvalg i hovedsak ekskludert. Dette er grunnen til at truede arter opplever det som kalles 'mutasjons-nedsmelting'. Av samme grunn kalkulerte Kimura størrelsen av sin ikke-seleksjons sone, som en enkel funksjon av populasjons-størrelsen (enten + eller - 1/2Ne). For genetikk-teoretikere er det meget attraktivt å begrense omfanget av støy til tilfeldige utvalg. Da kan en beleilig få støy til i hovedsak å forsvinne, ved enkelt å forutsette store populasjoner. Om det er behov for det kan også teoretikere få naturlig utvalg til å forsvinne ved å forutsette mye støy kombinert med flaskehalser i små populasjoner (eks. 'ut-fra-Afrika teorien').
Tilfeldig genetisk drift er meget sterk, og kan overstyre effekten av endog viktige mutasjoner i små populasjoner. I små populasjoner er naturlig utvalg i hovedsak ekskludert. Dette er grunnen til at truede arter opplever det som kalles 'mutasjons-nedsmelting'. Av samme grunn kalkulerte Kimura størrelsen av sin ikke-seleksjons sone, som en enkel funksjon av populasjons-størrelsen (enten + eller - 1/2Ne). For genetikk-teoretikere er det meget attraktivt å begrense omfanget av støy til tilfeldige utvalg. Da kan en beleilig få støy til i hovedsak å forsvinne, ved enkelt å forutsette store populasjoner. Om det er behov for det kan også teoretikere få naturlig utvalg til å forsvinne ved å forutsette mye støy kombinert med flaskehalser i små populasjoner (eks. 'ut-fra-Afrika teorien').  Begrensningene til naturlig seleksjon utvides ytterligere ut fra støy-problematikken. Støy-problematikken medfører at det meste av seleksjonens anstrengelser blir bortkastet. Det aktualiserer seleksjonskostnader og øker de reelle grensene som kostnad setter på naturlig seleksjon. Seleksjonsterskelen ved å selektere på for mange samtidige trekk, resulterer i fullstendig opphør av framgang. Ved å selektere etter ikke-arvelige faktorer sløses anstrengelsene bort. Det blir som prinsessa på erta har talløse erter på madrassen. Det vil gjøre problemet hennes verre. I anledning overlevelse av hele arter erkjente St. Jay Gould (yngre) betydningen av ulykker, overlevelse av den heldigste og støy. Men om støy rutinemessige overstyrer seleksjon, så blir langvarig evolusjon umulig. Det vil medføre genetisk degenerasjon og sluttelig artens utryddelse.
Begrensningene til naturlig seleksjon utvides ytterligere ut fra støy-problematikken. Støy-problematikken medfører at det meste av seleksjonens anstrengelser blir bortkastet. Det aktualiserer seleksjonskostnader og øker de reelle grensene som kostnad setter på naturlig seleksjon. Seleksjonsterskelen ved å selektere på for mange samtidige trekk, resulterer i fullstendig opphør av framgang. Ved å selektere etter ikke-arvelige faktorer sløses anstrengelsene bort. Det blir som prinsessa på erta har talløse erter på madrassen. Det vil gjøre problemet hennes verre. I anledning overlevelse av hele arter erkjente St. Jay Gould (yngre) betydningen av ulykker, overlevelse av den heldigste og støy. Men om støy rutinemessige overstyrer seleksjon, så blir langvarig evolusjon umulig. Det vil medføre genetisk degenerasjon og sluttelig artens utryddelse.Oversatt og tilpasset med figurer av Asbjørn E. Lund